Congratulations to Dr. Melanie Denommé Stauder for PhD graduation and Sophie Gauthier, BSc Honours graduation. We had spring/summer convocation last week and Melanie and Sophie were both there.
Melanie was the recipient of the Spirit of Brock award and a Distinguished graduate award. Here we celebrating outside my lab, although the photo-shy Sophie is not yet available:
Melanie saying goodbye to the lab and that’s me trying to convince her to stay!
Knowing how many animals are in a population sounds straightforward, but the method you use to count them can dramatically change the answer. Our new study, led by Patrick Moldowan as part of his PhD work at the University of Toronto, draws on 14 years of data from the long-running Bat Lake Integrated Salamander Study / Bat Lake Inventory of Spotted Salamanders (BLISS) at Algonquin Provincial Park to put three common amphibian monitoring approaches head-to-head: aquatic funnel trapping, egg mass counting, and coverboard surveys (Figure 1). By comparing all three against a kilometre-long drift fence encircling the entire lake and intercepted nearly every salamander on its breeding migration, we were able to ask which methods actually give reliable estimates of population size and sex ratio for spotted salamanders (Ambystoma maculatum). The drift fence revealed annual breeding populations of between 4,500 and 5,300 individuals, a strikingly large population that reflects the intact, fish-free nature of Bat Lake.
Figure 1. Salamander monitoring methods at Bat Lake, Algonquin Provincial Park, Ontario, Canada: aquatic funnel trapping (2008–2019), drift fence with terrestrial funnel traps (2017–2019), and coverboards (2019–2020). Egg masses were counted throughout the annual breeding period (2008–2019) by paddling the vegetated perimeter of the lake.
The results carry some important cautionary lessons for wildlife monitoring. Funnel traps alone underestimated population size by 67–75%, largely because individuals differ enormously in how likely they are to be caught. When this individual heterogeneity in capture probability was accounted for in the models, estimates aligned much more closely with the fence counts. Even more striking was what the traps suggested about sex ratios: raw trap data implied males outnumbered females two-to-four-to-one, when in reality the population was slightly female-biased. Coverboards, meanwhile, detected less than 1% of the population and were essentially uninformative for this species. Egg mass counts, often used as a convenient proxy for female abundance, were only weakly correlated with actual female numbers at this site, showing that these monitoring shortcuts need to be validated before being relied upon. Together, these findings offer practical guidance for anyone designing long-term amphibian monitoring programs, emphasizing that how you count is just as important as what you count.
Citation
Moldowan, PD, Armstrong, DP, Tattersall, GJ, and Rollinson, N. 2026. A comparison of monitoring methods for inferring demographics of a pond-breeding amphibian population over 14 years. Journal of Wildlife Management, e70225. https://doi.org/10.1002/jwmg.70225
I am really proud to congratulate Dr. Melanie Denommé Stauder for her successful PhD Defence!
The thesis entitled “Analyzing common husbandry practises and potential indicators of welfare in Pogona vitticeps” was defended today in front of her examining committee: Dr. Noam Miller (External, Wilfrid Laurier University), Dr. Dustin Duncan (External within Brock University), Dr. Michael Owen (Chair, Brock University), Dr. Georgia Mason (University of Guelph), Dr. Gaynor Spencer, and myself.
Great discussions were had and some new aphorisms learned. “We are not reptiles”, and “we are never as smart as we need to be” are some gems of wisdom to take away from the defence.
Despite being a major event in early tetrapod evolution, many living fish species occasionally leave water, but the reasons are not always obvious. Some do it to feed, some to escape predation, and some to cope with poor water quality, although rarely for long periods of time. Another intriguing possibility is that fish may use land to help regulate their body temperature.
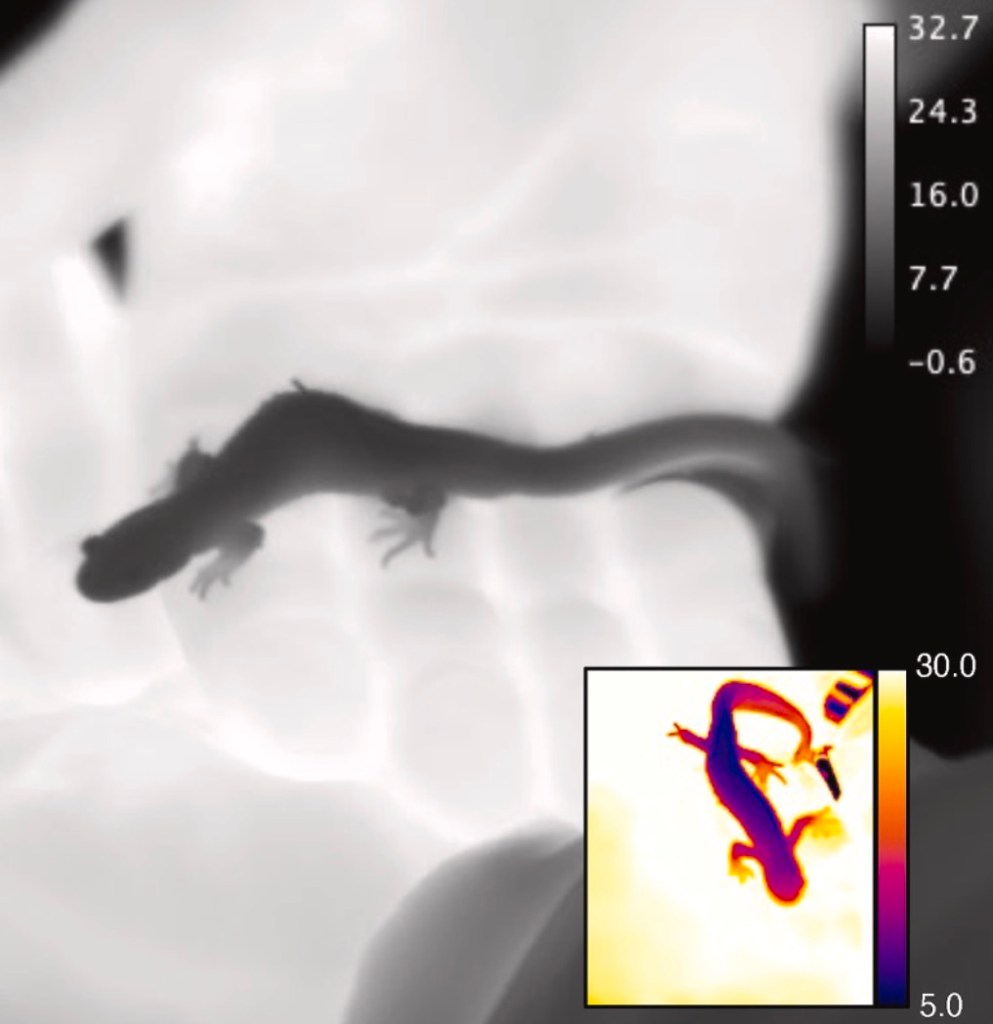
The mangrove rivulus (Kryptolebias marmoratus) is a particularly good species to explore this question. This small tropical fish is famous for its ability to survive on land for weeks at a time, provided the humidity levels are high. When water gets warm, mangrove rivulus will often emerge onto land, where evaporative cooling can rapidly lower body temperature, as we showed previously (https://doi.org/10.1098/rsbl.2015.0689). But leaving the water also comes at a cost: gas exchange becomes much more difficult, leading to temporary oxygen limitation and CO₂ buildup.
The mangrove rivulus fish (Kryptolebias marmoratus), is typically found in areas with Red Mangrove trees, like Florida, Mexico and Brazil. (Photo from Dr. Andy Turko)
In many ectotherms, low oxygen levels are known to shift thermal preferences toward cooler temperatures. That led us to ask a simple question: when rivulus go onto land, do they actively seek cooler temperatures than they would in water?
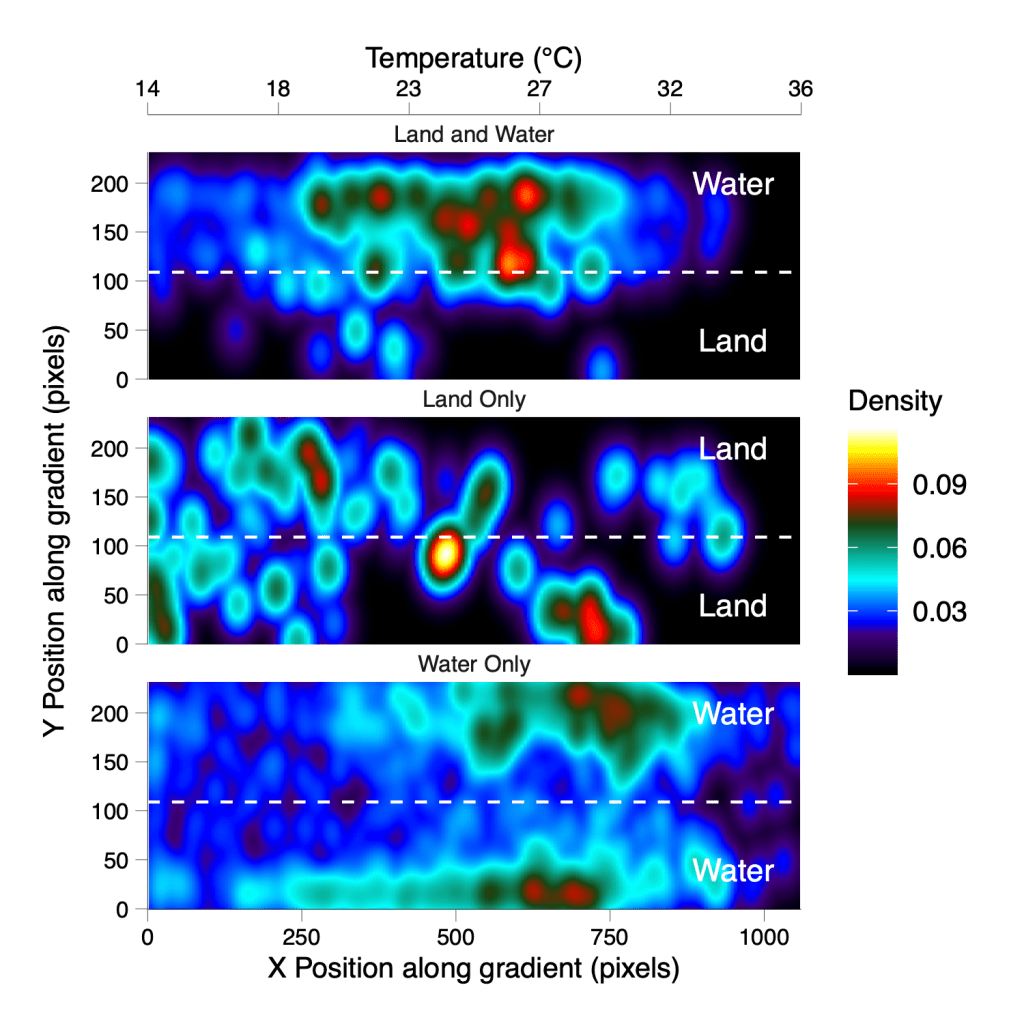
To test this, we (former Honours students Katlyn Dundas and Philip Bartel) gave fish access to choice of temperatures along a thermal gradient under three conditions: water only, land only, or a combination of both. What we found was striking. Rivulus only selected cooler temperatures when they were on land. When confined to water, their preferred temperatures remained higher, even when the same thermal options were available.
This suggests that their terrestrial emergence is not simply a passive escape from warm water to benefit from evaporative cooling. Instead, fish on land appear to actively choose cooler environments, which is a response consistent with anapyrexia, the deliberate reduction in the regulation of body temperatures (analogous but quite different from fever). In this case, leaving the water may help offset the physiological challenges of breathing air through gills that are adapted for water.
Together, these results add a new piece to the puzzle of why some fishes venture onto land. For mangrove rivulus, emerging from water may provide more than just temporary cooling. It may fundamentally change how they regulate temperature when oxygen becomes limiting.
The study is now available at the Journal of Experimental Biology website, citation below.
Citation
Dundas, KE, Bartel, PB, Turko, AJ, and Tattersall, GJ. 2026. Terrestrial emergence reflects lower thermal preferences in the mangrove rivulus (Kryptolebias marmoratus). Journal of Experimental Biology, 229 (4): jeb251829. https://doi.org/10.1242/jeb.251829
Many thanks to Brock University Library open access fund for supporting publication in the Journal of Experimental Biology!
Smooth scatter plot of mangrove rivulus space use within the thermal gradient chamber and depicting the three conditions tested.
Measuring body temperature should seem straightforward, but in ectotherms, the act of measuring can be part of the problem. For especially small amphibians in particular, common approaches like cloacal thermometry require restraint and direct contact, raising the possibility that handling itself alters body temperature before it is even recorded. In this study, we asked a simple but important question: does brief, gentle handling measurably change the body temperature of salamanders?
Using infrared thermography, we examined how short periods of handling affected skin temperature in two closely related mole salamanders, the blue-spotted salamander (Ambystoma laterale) and the spotted salamander (A. maculatum). We also combined field measurements with controlled laboratory experiments to tease apart the effects of physical contact, heat transfer, and behaviour.
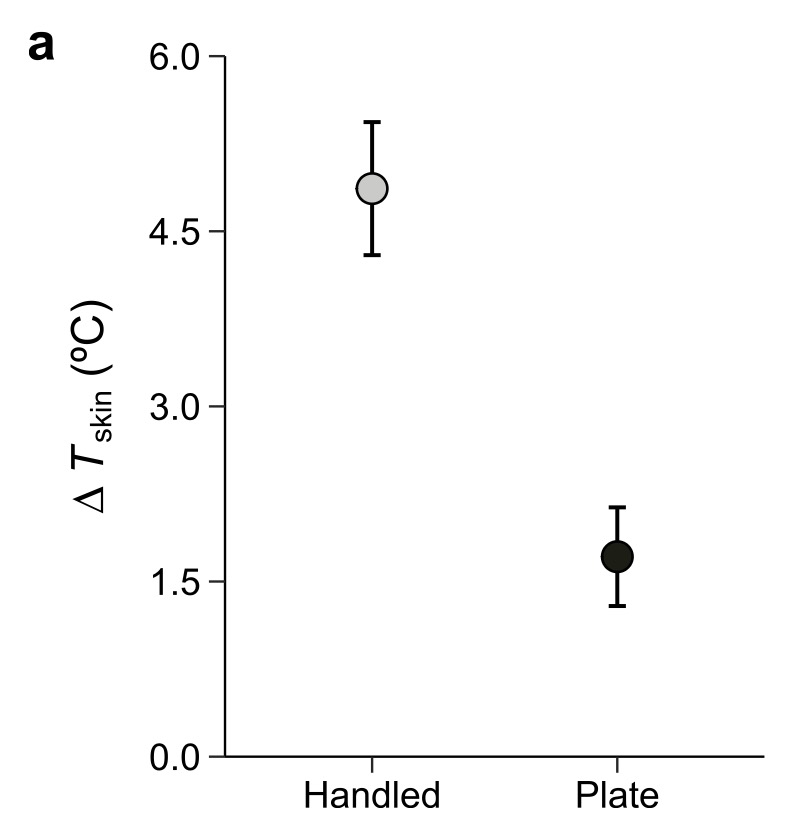
The answer was clear. Even short handling periods caused salamanders to warm rapidly. In the field, handled animals were consistently warmer than unhandled controls, with the head warming more than the rest of the body. Not surprisingly, smaller salamanders showed larger temperature changes, suggesting body size plays an important role in how quickly heat is gained.
Change in skin temperature of yellow spotted salamanders either handled or placed on a warm plate (set to the same temperature as human handler).
In the lab, salamanders placed on a warm surface (set to hand temperature) also warmed up, but not as much as animals that were actually handled. This tells us that handling is not simply about contact with something warm. While we don’t know precisely why yet, the most likely explanation is that salamanders manifest a stronger cardiovascular response to human handling and the heat that they pick up is transferred more quickly throughout the body.
Temperature changes matter from a behavioural perspective. Warmer salamanders were more likely to become active, regardless of whether that warmth came from handling or from contact with a warm surface. That means handling can influence not only the temperature we measure, but also the behaviour we observe. Taken together, these results highlight a subtle but important issue: brief handling can rapidly alter body temperature, sometimes at rates comparable to experimental heat stress studies. For researchers, this has implications for data accuracy in thermal biology. More broadly, it is a reminder that even well-intentioned, gentle handling can have unintended physiological effects, especially for small ectotherms.
Some caveats: this study is not to be interpreted as alarmist. Warming up from being in contact with warm temperatures is a pretty obvious expectation. These results would only have possible impacts for studies concerned with accurate temperature measurements or those that perform very brief behavioural observations during or immediately after handling (which is probably very rare) that are temperature dependent. And as one of the reviewers of the manuscript rightly pointed out, we measured surface temperatures, which will differ and lag with core temperature measurements.
We do note that there is a related study appearing in the same issue of the Journal of Thermal Biology highlighting similar cautions as this one, and that particular study actually measures core temperature of really tiny frogs!
Citation
Giacometti, D, Montes, LF, Denommé, M, Andrade, DV, and Tattersall, GJ. Handle with care: the thermal consequences of short-term handling in mole salamanders. Journal of Thermal Biology, 136: 104390. https://doi.org/10.1016/j.jtherbio.2026.104390
A lot of news has happened in the past month and I have been negligent about updating. All lab members have had awards, scholarships, and bursary announcements that warrant celebration!
• Melane Denommé Stauder (PhD student) will be receiving the Jack Miller Graduate Award for the Faculty of Mathematics and Sciences this coming year. Stay tuned for a future announcement as I’m not sure this is public knowledge yet.
• Harry Kumbhani received the Ontario Paper Thorold Foundation Graduate Award, in recognition of academic standing and that his eco-physiological research is aligned with environmental research.
• Harry Kumbhani also was lead author on a paper published in Canadian Journal of Zoology that was highlighted as Editor’s choice.
• Danilo Giacometti’s (recent PhD graduate) Journal of Experimental Biology paper from 2025 was a finalist for the JEB Outstanding Paper Prize. Link still to come.
• Sophie Gauthier (Biological Sciences Honours Thesis Student in my lab) was awarded the Niagara Birds Bursary for her work in an ecologically relevant project (awarded to a Biological Sciences student engaged in a research project in ecology or related discipline.
Congratulations to all and a huge thanks to those who have supported student and ECR success through the funding of these scholarships and establishment of these recognitions.
Danilo Giacometti was interviewed by NPR a couple of weeks ago about our blue-spotted salamander paper in CJZ, and the interview came out today (December 24, 2025). Check it out here:
A recent paper from the lab, led by PhD student Melanie Denommé, examines a foundational assumption in reptile husbandry: that animals will inherently prefer naturalistic enclosures over simpler, more utilitarian ones. Using bearded dragons (Pogona vitticeps) as a model species, Melanie conducted preference tests in which lizards were given free choice between naturalistic and standard enclosure styles (Figure 1). Critically however, individuals had been reared under two different housing conditions and tested at multiple time points across their lives, allowing us to shed light on preferences shaped by experience. Contrary to expectations, the dragons did not show an immediate or universal preference for the more naturalistic enclosures. Instead, their choices were strongly influenced by prior housing conditions, age, and timing of the tests, demonstrating that familiarity and developmental context play an important role in how captive reptiles evaluate their environments.
Figure 1. Graphic abstract from the study, describing the timing of the preference tests and housing conditions lizards were under at the two time points when preferences were tested. The y-axis describes a bias index, describing what proportion of time lizards spent within each enclosure style (values of 0 indicated 100% of time spent in standard, values of 1 indicate 100% of time spent in naturalistic).
Despite the absence of a consistent enclosure-level preference, the behavioural data told a more nuanced and compelling story. When lizards engaged in key species-typical behaviours such as climbing, digging, and hiding, they overwhelmingly used naturalistic resources rather than the simpler substitutes provided in standard enclosures. This finding suggests that while reptiles may not always “choose” a naturalistic enclosure outright, these environments are clearly superior for facilitating motivated behaviours that are closely linked to welfare. As part of Melanie’s broader PhD research program, this work emphasizes the importance of empirically testing husbandry assumptions rather than relying only on tradition or aesthetics alone, and it provides concrete guidance on which enclosure features matter most from the animal’s perspective. More than that, we argue that context and experience are important considerations in these kinds of studies.
Citation
Denommé, M and Tattersall, GJ. 2026. Try before you buy: Preferences for naturalistic-style enclosures are influenced by experience in bearded dragons (Pogona vitticeps). Journal of Applied Animal Behaviour Science, 295:106887. https://doi.org/10.1016/j.applanim.2025.106887
Our new paper, led by MSc student Harry Kumbhani and building on fieldwork conducted by former MSc student Curtis Abney, explores how Eastern Garter Snakes (Figure 1) navigate the complex thermal landscapes of wetlands in southern Ontario. Using detailed operative temperature models, which were copper snake replicas equipped with temperature loggers, our team quantified how three adjacent habitat types (closed forest, mixed shrub, and open peat; Figure 2) differ in the thermal opportunities they provide. Although the open peat habitat consistently reached the warmest temperatures and offered the greatest access to the snakes’ preferred thermal range, it also exhibited extreme highs that frequently exceeded the species’ upper thermal tolerance. This created a paradox: the habitat with the highest apparent thermal quality was also the riskiest.
Figure 1. Eastern garter snake (Thamnophis sirtalis sirtalis).Figure 2. A snake’s eye view of the canopy cover within the three habitat types.
Despite expectations that snakes might favour the warmest habitat, we found that Eastern Garter Snakes were most abundant in the intermediate, mixed-shrub habitat, a pattern we describe as a “Goldilocks effect.” This middle habitat provided a balance of sun and shade, offering both basking opportunities and safe retreat sites, and avoided the thermal instability and overheating risk found in the open peat. The study suggests that thermal quality is more nuanced than simply being warm enough; stability, heterogeneity, and safety from extreme temperatures all shape how snakes use their environment. These findings highlight the importance of structurally diverse habitats for temperate reptiles and offer valuable insights into how changing landscapes may influence thermoregulation and habitat choice in the future.
Citation
Kumbhani, HAW, Abney, CR, Giacometti, D, and Tattersall, GJ. 2025. Operative temperatures of Eastern Garter Snakes (Thamnophis sirtalis sirtalis) reveal a Goldilocks effect for habitat use. Canadian Journal of Zoology, 103: 1-15. https://doi.org/10.1139/cjz-2025-0090
A new study led by Morgane Vandendoren, Nicole Bedford, and others from Adam Nelson’s lab at the University of Wyoming has uncovered a new role for oxytocin, the so-called “love hormone.” Published in eLife, the research shows that oxytocin neurons in the paraventricular hypothalamus act as a kind of biological switch, helping mammals transition from cooler, resting states to warmer, active ones. Using mice, the team combined calcium imaging, optogenetics, and behavioural observation to show that bursts of oxytocin neuron activity reliably occur just before an animal warms up and becomes active, even in the absence of social cues. These neurons appear to coordinate both thermogenic (heat-producing) and behavioral arousal, revealing a new layer of oxytocin’s influence that bridges physiology and behaviour.
This collaboration, with contributions from several Wyoming students and collaborators, demonstrates how oxytocin not only shapes social and maternal behaviours but also tunes the body’s thermal and arousal cycles. My lab’s involvement was a bit on the periphery, but focused on the thermal imaging and coding pipelines that helped visualize these rapid transitions in body temperature and activity. Together, the findings expand our understanding of oxytocin beyond its traditional social context, showing that it also plays a key role in the daily rhythm of energy balance and physiological readiness.
The University of Wyoming have a more detailed press release for the study here:
This paper was published in eLife, following an open peer review approach that I am still trying to wrap my head around. The citation is below, and so formally the study is published in preprint format, with us having still to upload a revised manuscript which will address some of the points raised by the reviewers.
Citation
Vandendoren, M, Rogers, JF, Landen, JG, Killmer, S, Alimiri, B, Pohlman, C, Tattersall, GJ, Bedford, NL, Nelson, AC. 2025. Oxytocin neurons signal state-dependent transitions to thermogenesis and behavioral arousal in social and non-social settings. eLife, 14: RP108212. https://doi.org/10.7554/eLife.108212.1