We have been studying behavioural thermoregulation in various ectotherms for years. Unlike what is commonly taught, ectothermic animals (e.g. amphibians, reptiles, fish, invertebrates) can and do respond to variation in temperature in their environment, and are not completely helpless in the face of thermal variation. Many choose specific temperatures, bask in the sun, avoid thermal extremes, or have evolved strategies to take advantage of predictable thermal variation in the environment. In other words, many exhibit preferred temperatures. We can study this in the lab in a number of ways. The first is a thermal gradient (see the banner image above). This simple approach allows animals to indicate thermal preference by simple movement choices.
A second, more convincing approach is through the use of a shuttle box. This concept relies on a form of operant conditioning, where animals must choose a chamber corresponding to the more comfortable temperature. Two chambers are connected by a simple doorway, one chamber is warmer than the other. More sophisticated shuttle boxes incorporate ra thermal change over time, such that decisions the animal makes has consequences for the future temperature.
Choosing the warm side indicates an exit temperature that can be scored. After entering the warm side, both chambers slowly rise in temperature, unless the animal escapes back to the cool chamber. The process repeats, except that both chambers automatically cool. Over time, you end up with a series of exit temperatures that should reflect the tolerable, preferred, or “comfortable” range of temperatures. We have successfully studied this is fish, lizards, and crayfish.
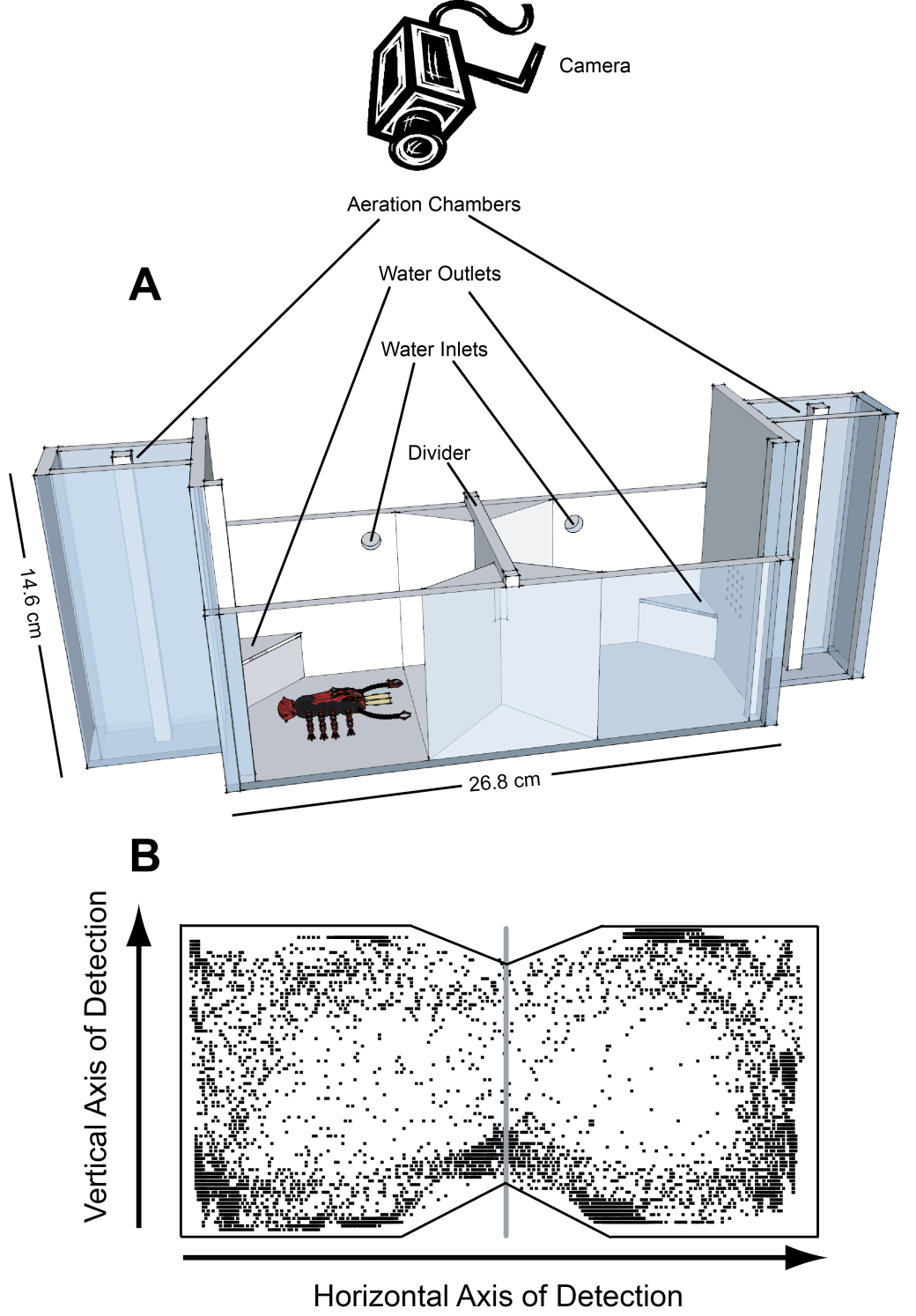
Custom built thermal shuttle box chamber and webcam detection system for the study of behavioural thermoregulation, thermosensation, thermal avoidance behaviours in aquatic organisms.
We have also examined remote temperature sensing behaviour in the ectothermic rattlesnake. These animals have highly sensitive thermal pit organs in the rostral area that are used for sensing warm-blooded prey, as well as orienting for thermoregulatory behaviour.
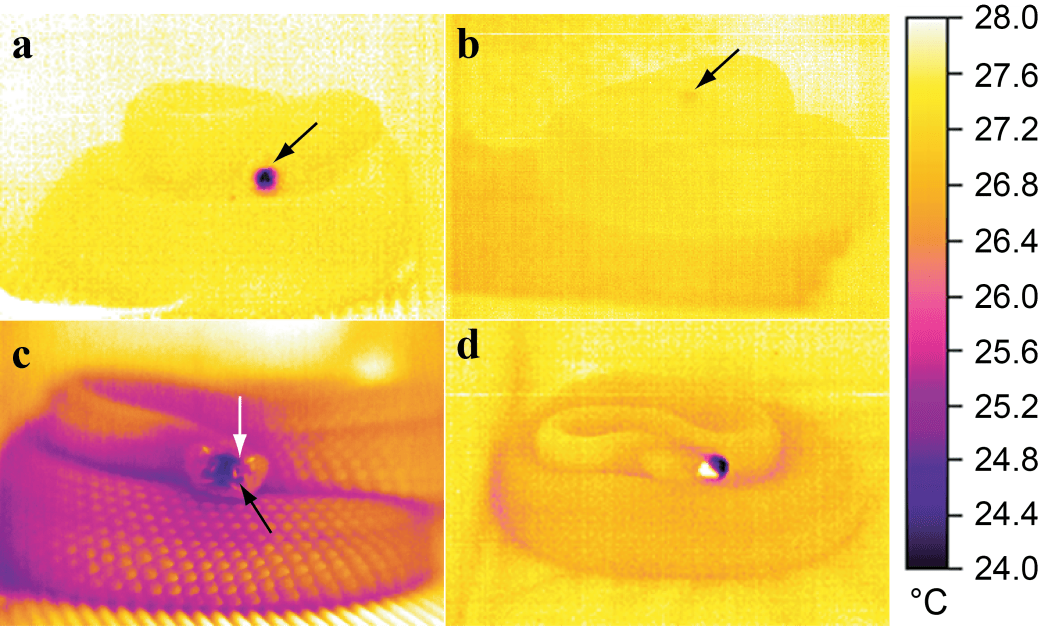
Infrared thermal images of a South American rattlesnake under low (a) and high humidity (b) conditions, demonstrating how breathing forces substantial cooling of the face and pit organ region (c).
Source Material
Cadena, V, Andrade, DV, Bovo, RP, and Tattersall, GJ. 2013. Evaporative respiratory cooling augments pit organ thermal detection in rattlesnakes. Journal of Comparative Physiology A 199: 1093-1104.
Tattersall, GJ, Luebbert, JP, LePine, OK, Ormerod, KG, and AJ Mercier. 2012. Thermal games in crayfish depend on establishment of social hierarchies. Journal of Experimental Biology 214: 1892-1904.
Tattersall, GJ, Tyson, TM, Lenchyshyn, JR, and RL Carlone. 2012. Temperature preference during forelimb regeneration in the red-spotted newt Notophthalmus viridescens. Journal of Experimental Zoology Part A Ecological Genetics and Physiology 318: 248-257.
Khan, JJ, Richardson, JML, and Tattersall, GJ. 2010. Thermoregulation and aggregation in neonatal bearded dragons (Pogona vitticeps). Physiology & Behavior. 100: 180-186.
Cadena, V and Tattersall, GJ. 2009. Decreased thermoregulatory precision contributes to the hypoxic thermoregulatory response in lizards. Journal of Experimental Biology. 212: 137-144.
Cadena, V and Tattersall, GJ. 2009. The effect of thermal quality on the thermoregulatory behaviour of the bearded dragon, Pogona vitticeps: influences of methodological assessment. Physiological and Biochemical Zoology, 82: 203-217
Tattersall, G. J. and Boutilier, R. G. 1999. Does behavioural hypothermia promote exercise recovery in cold-submerged frogs? Journal of Experimental Biology, 202: 609-622.
Tattersall, G. J. and Boutilier, R. G. 1997. Balancing hypoxia and hypothermia in cold-submerged frogs. Journal of Experimental Biology 200: 1031-1038.
