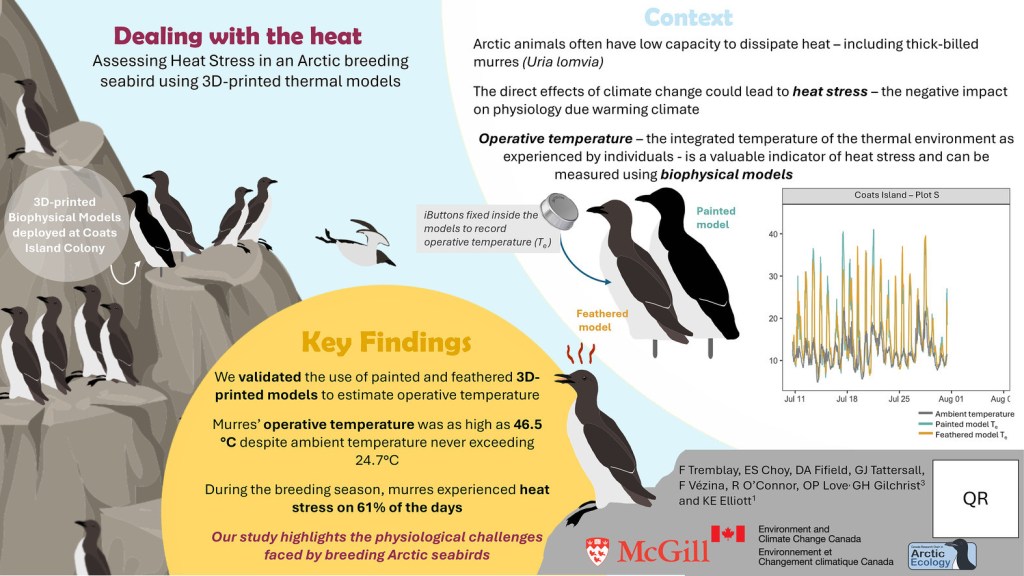
In the cold, temperate forests, long before spring fully arrives, blue-spotted salamanders (Ambystoma laterale) are already on the move. These small amphibians begin migrating to their breeding ponds while snow still blankets the ground and ice lingers on and in the soil. This is a risky strategy for a species that can’t survive freezing. Our recent Natural History note, spearheaded by Dr. Danilo Giacometti and published in the Canadian Journal of Zoology, documents this remarkable early migration and presents new thermal imaging evidence that blue-spotted salamanders achieve this while at sub-zero body temperatures.

For amphibians like blue-spotted salamanders, freezing is typically fatal. Ice crystals rupture cells, leading to irreversible damage. Unlike some frogs that survive being partially frozen thanks to natural antifreezes like glucose, blue-spotted salamanders are known to be freeze-intolerant.
But in spring 2022 in Algonquin Park, during a brief window of opportunity we observed salamanders actively migrating, even while walking across or sheltering beside ice. Using high-resolution thermal imaging, we measured their skin temperatures (a reliable proxy for body temperature in such small animals) and found several individuals with body temperatures as low as –3.6°C, which is well below their known freezing point. Our findings suggest that blue-spotted salamanders may rely on supercooling, where their body fluids remain liquid even below freezing. This strategy has been shown in lab studies to be possible down to about –1.5°C, but our field data suggest some individuals may supercool even further, albeit briefly.
One of the most surprising observations was that several salamanders were in direct contact with ice, a known trigger for freezing of fluids that are supercooled. Despite this, they were active and moving, raising fascinating questions about how they might avoid nucleation (the start of ice formation) in natural settings or if they can manage short-term freeze/thaw during their migration.

Why would salamanders take such a risk by migrating so early? There may be several evolutionary advantages. By arriving at breeding ponds early before other species, they reduce competition and potentially avoid predators. Early breeding also gives their offspring more time to grow before winter returns.
Our study opens new questions about the limits of amphibian cold tolerance and the role of behavior and microhabitat selection. More research is needed to understand whether these salamanders truly remain supercooled for long periods or whether they occasionally freeze and recover, a possibility hinted at but not yet proven in this species.
For now, our thermal images offer a rare glimpse into the early spring lives of blue-spotted salamanders and reveal that there’s still much to learn about how animals survive the cold.
You can access our open data and images here: https://doi.org/10.5683/SP3/FZJBQH and the paper at the following doi:
Citation
Giacometti, D, Moldowan, P, and Tattersall, GJ. 2025. Sub-zero body temperatures during early spring migration in blue-spotted salamanders (Ambystoma laterale). Canadian Journal of Zoology, https://doi.org/10.1139/cjz-2025-0045
Postscript: An Editor’s Lament
The journey that this very brief natural history note took to reach publication was unnecessarily arduous. We originally submitted this study to Canadian Field Naturalist in August 2022. In that initial submission, we heard back after 16 months from the editor that the manuscript had been peer reviewed (3 reviewers) and with straightforward revisions; we supplied revisions within 30 days in January 2024. Then all went quiet with the journal for months. We reached out on numerous occasions to the editors in 2024 about whether the manuscript was still being handled, whether we would hear a decision, and received responses that indicated that editing it was not a high priority.
So, after 2.5 years sitting with Canadian Field Naturalist, we withdrew the manuscript (Spring 2025) and submitted it to CJZ where I am pleased to note that the manuscript underwent a normal peer review process.
As an editor (the average turnaround time for 1st submit Major/Minor decision papers I handle is 51 days – this includes the time to find reviewers), I was saddened at how CFN handled the initial manuscript. There were extenuating circumstances in that the associate editor handling the initial submission passed away, but we were assured by the editorial team that the manuscript would not get ‘lost’ in the re-shuffle.
As a society journal, it deserves support, but 2.5 years to handle a short manuscript does not set a good example for early career researchers; this is almost a lifetime for a graduate student.
I understand that editors need to make difficult decisions and in the course of those duties often reject studies (for fit or for other reasons). But timely decision making is just as important or more so for early career researchers. All the academic editors I know are full-time employed with academic and research jobs, but I have never heard any of them indicate or hint to an author that their submission is not a priority. If the work is not appropriate for the journal, the most humane decision is to reject it in a timely manner.
Anyhow, I am pleased with the Canadian Journal of Zoology’s handling of the manuscript. It was professional and straightforward and now we can move on from this experience.
I do think that we need to support natural history style studies/observations, so I can only hope that by sharing this, those that read this may push for change at journals that could use the support.